
На протяжении тысяч лет человечество стремилось в небеса, желая уподобиться птицам. Многие великие умы, от Леонардо да Винчи до братьев Райт, проектировали самые разнообразные летательные аппараты, одни из которых так и остались на бумаге, а другие стали прародителями современных самолетов и вертолетов. Сейчас полет для человека не составляет труда — купил билет на самолет и вуаля. Большинство из нас даже не задумывается о том, как работает эта огромная металлическая птица, способная перенести нас с одного континента на другой. Правда между современными самолетами и реальными птицами общего не так и много, но с разрабатываемыми автономными роботами ситуация немного иная. Многие ученые пытаются создать робота-птицу, а дабы это начинание было успешным, необходимо понимать не только как птицы летают, но и как приземляются. Сегодня мы с вами познакомимся с исследованием, в котором выпускники Стэнфордского университета подробно изучили механизм посадки попугайчика по имени Гэри и его сородичей. Как птицы приземляются, как они понимают, что могут или не могут приземлиться на той или иной поверхности, и какова кинематика их конечностей во время посадки? На эти и другие вопросы мы найдем ответы в докладе исследователей. Поехали.
Основа исследования
Чем отличаются современные летательные аппараты от птиц в аспекте посадки? Боингу 777, например, нужна взлетно-посадочная полоса длиной минимум 2500 м, вертолету нужна ровная (желательно) поверхность, как и коптерам. Птицы, в свою очередь, не ограничены в геометрии посадочного полотна и могут приземлиться практически на любой поверхности (если рассматривать птиц в целом, а не отдельно взятый вид). Если же говорить об автономных роботах, то их возможности в плане посадки весьма ограничены. Некоторые из них могут успешно садиться на специально спроектированные поверхности, но это не идет ни в какое сравнение с обыкновенным дятлом, который может приземлиться на вертикальной, наклонной, горизонтальной поверхностях, лишь бы было за что коготками зацепиться.

Мне и так удобно (муравьиный дятел / Melanerpes formicivorus).
Исследователи отмечают, что их предшественники уже проводили некоторые исследования, которые дали количественную оценку динамики ног и крыльев птиц во время посадки. Однако остается неясным, как эта динамика меняется в зависимости от разного типа поверхностей, служащих местом посадки. Ведь важна не только геометрия области посадки, но и текстура поверхности.
Любопытно еще и то, что предыдущие исследования проводились не на живых птицах, следовательно, полноценно оценить силу захвата лапок и когтей на практике не представлялось возможным. Оттого и остались неосвещенными некоторые вопросы: как именно птицы понимают, что место посадки подходящее; какая динамика конечностей в момент касания и захвата; почему для птиц посадка на самые разные поверхности это плевое дело и т.д.
В рассматриваемом нами сегодня труде исследователи решили проверить механизмы и динамику приземления древесных птиц, проведя наблюдения за воробьиными попугайчиками Лессона (Forpus coelestis).

Воробьиный попугайчик Лессона.
Подопытные добровольно приземлялись на жердочки с различной геометрией и текстурой. Всего было 9 вариантов: 3 «естественных», 3 с измененным диаметром и 3 с измененной текстурой.
«Естественные» варианты названы так, поскольку жердочки были сделаны из настоящих веточек (19 мм в диаметре) трех разных деревьев:
- дуб калифорнийский — поверхность шероховатая в разных участках одной ветки;
- сейба великолепная — поверхность относительно гладкая, ветки мягкие;
- османтус душистый — поверхность продольно шершавая, ветки жесткие.
Для опытов с измененным диаметром жердочки в качестве материала были использованы обычные березовые нагели разного диаметра:
- 38 мм — слишком большой диаметр для данного вида попугаев;
- 19 мм — попугай может выполнить частичный захват (передние и задние пальцы не соприкасаются);
- 6 мм — попугай может выполнить полный захват (передние и задние пальцы соприкасаются).
Для опытов с разными текстурами березовую жердочку диаметром 19 мм обернули в три разных материала:
- пена — мягкая и упругая текстура;
- тефлон — скользкая текстура;
- наждачная бумага — грубая и шероховатая текстура.
Попугайчик тестирует все 9 вариантов посадочной площадки.
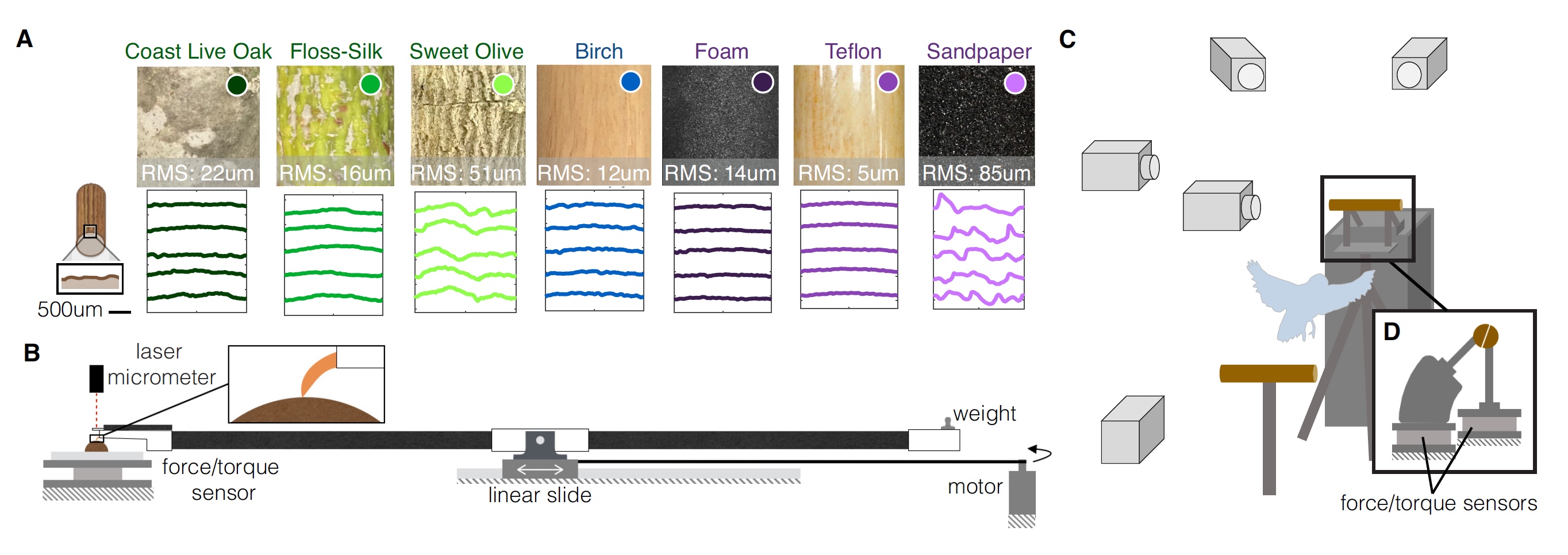
Изображение №1
Было проведено трехмерное структурно-световое сканирование каждого из вариантов жердочки, что позволило количественно описать текстуру поверхности каждой из них и воссоздать профили поверхности (1А). Чтобы оценить влияние этих текстурных особенностей на взаимодействие поверхности жердочки и стопы попугая, исследователи провели тесты сопротивления пальцев и когтей, чтобы измерить силы трения для каждой поверхности. Также была оценена и сила давления когтей, чтобы измерить деформацию поверхности (1B).
На схеме 1С мы видим, что все тесты проводились под пристальным взглядом нескольких скоростных камер, а жердочки были распилены пополам. К каждой половинке присоединен датчики силы и момента силы.
Результаты исследования
Прежде всего рассмотрим динамику крыльев и лапок во время посадки.
Во всех тестах со всеми вариантами посадочной поверхности подопытные демонстрировали одинаковую классическую динамику. Посадка начинается с торможения крыльями (воздушная фаза), после чего лапки поглощают оставшийся импульс при контакте с жердочкой (фаза поглощения). Контакт осуществлялся обеими лапками с временной задержкой между ними в несколько миллисекунд, т.е. первой с жердочкой контактировала предпочтительная лапка:
- подопытный №1 — в 100% случаев правая лапка;
- подопытный №2 — в 83% случаев левая лапка;
- подопытный №3 — в 85% случаев левая лапка.
После контакта наступает фаза закрепления, когда попугайчик более надежно и крепко фиксируют пальцы вокруг жердочки. Следом следует этап, которые встречается не всегда, но часто, — фаза корректировки, когда они тем или иным образом меняют положение лапок или пальцев на жердочке.
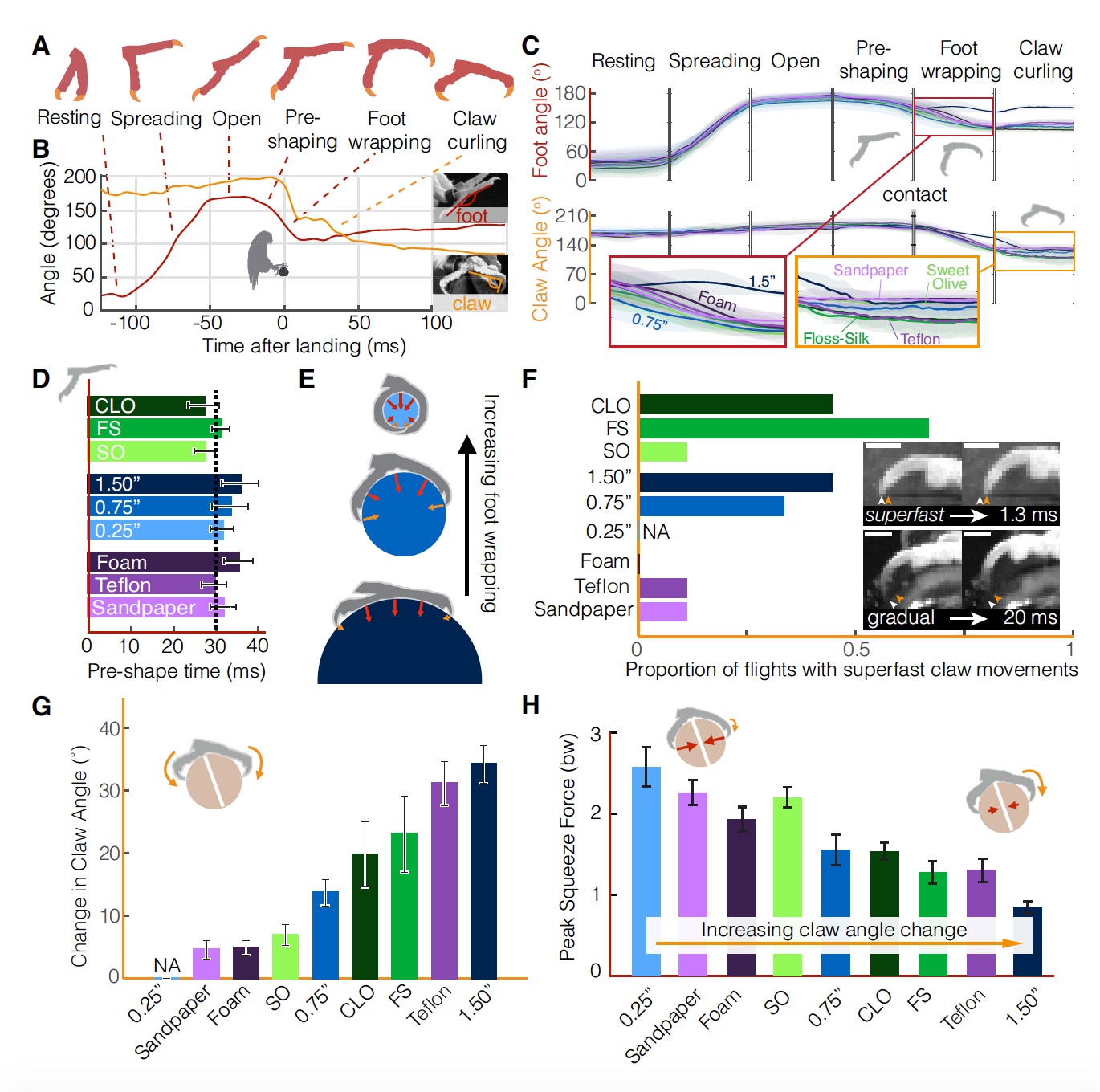
Все фазы посадки попугая (соответствует схеме на 2А).

Изображение №2
Иногда попугаи, видимо переоценив или недооценив условия теста, немного недолетали/перелетали место посадки (2В), что приводило к большей дисперсии угла наклона стопы, при котором они устанавливают статическое сцепление.
Любопытно, что при наличии некоторой вариативности в показателях силы лапок (2В) и направления (2C) средние значения силы посадки остаются удивительно одинаковыми для разных жердочек.
Различие показателей, замеченные во время приземления, может быть объяснено стратегией приземления попугаев. Есть теория, в соответствии с которой птицы контролируют свою посадку, визуально оценивая время контакта с поверхностью, τ(t). Для успешного приземления они регулируют свою скорость захода на посадку, чтобы поддерживать постоянную τ(t). Если немного детальнее: τ определяется как расстояние до жердочки (s), деленное на скорость приближения (v).
Если птица тормозит с постоянным замедлением (а), то τ = s/v = 0.5at2/at = 0.5t, в таком случае τ(t) = 0.5.
Если τ(t) τ(t)
Учитывая вышеописанную кинематику попугаи поддерживают относительно постоянные значения τ (2Е), соответствующие значениям контролируемых столкновений (2F). Самое малое среднее значение τ = 0.80 наблюдалось в тестах с жердочкой с наименьшим диаметром (6 мм). А самое большое значение τ = 0.94 было в тестах с жердочкой, покрытой пеной. Это говорит о том, что попугаи увеличивают торможение, когда приближаются к мягкой посадочной поверхности.
Наличие высокого показателя τ (т.е. больше 0.5) во всех тестах со всеми типами поверхности говорит о том, что в момент касания птицы еще не завершили посадку.
Немного ознакомившись с динамикой посадки, перейдем к кинематике стоп и когтей наших пернатых друзей.
Учитывая однотипность поведения во время посадки вне зависимости от типа поверхности, исследователи сделали вывод, что птицы реагируют на различия в текстуре и геометрии посадочной жердочки именно лапками и когтями, а не крыльями. Звучит логично, не правда ли? Но, сюрприз, это не совсем так.
Кинематика стопы продемонстрировала сходный стереотипный (одинаковый для всех типов жердочек) набор посадочных этапов (3A, 3B).
Изображение №3
Во время полета стопы находятся в закрытом состоянии (фаза отдыха), далее они начинают открываться примерно за 100 мс до касания с жердочкой (фаза раскрытия длительностью 40 ± 8 мс), далее идет «открытая» фаза, когда в течение 21 ± 7 мс стопа полностью раскрыта.
Непосредственно перед контактом пальцы начинают сближаться (фаза предварительного формирования захвата, длительность 31 ± 10 мс). После установления контакта с поверхностью пальцы начинают обхват жердочки (фаза обхвата, длительность 19 ± 7 мс). Заключительная фаза это захват когтями длительностью 185 ± 11 мс.
Различия в текстуре и геометрии жердочки влияют исключительно на кинематику этапов посадки после контакта с поверхностью (3С). До соприкосновения кинематика стопы и когтя остается неизменной во всех тестах, даже когда стопы начинают закрываться во время фазы предварительного формирования захвата.
Другими словами, если бы птицы реагировали на разницу в геометрии и текстуре посадочной поверхности, то изменения кинематики наблюдались бы еще во время фаз до контакта, т.е. еще в воздухе. Вместо этого попугаи последовательно начинают предварительное формирование захвата за 30 мс до приземления (3D). Это наталкивает на предположение, что у них есть прогнозируемые варианты решения задачи по посадке на необычной поверхности.
Для птицы важно сохранять большой угол между когтями до самого момента контакта, чтобы предотвратить их повреждение. Далее степень захвата определяется диаметром жердочки (3Е). Полный захват происходит в случае жердочек диаметром 6 мм, т.е. попугаи могут полностью использовать всю свою силу пальцев и когтей для стабилизации.
Важно отметить, что в момент контакта когти и/или лапки могут соскальзывать с жердочек, ввиду гладкой поверхности. В результате угол захвата когтя может изменяться. Принимая во внимание, что стадия захвата когтями может длиться более 100 мс, отдельно взятые движения отдельно взятого когтя (3F) могут происходить невероятно быстро (1-2 мс).
Сверхскоростные движения когтями, вероятно, не регулируются только мышечным сокращением, как предполагают исследователи. Удивительно и то, что среди позвоночных самые быстрые движения мышц в 5-10 раз медленнее, чем эти супербыстрые движения когтей (например, мышцы рук человека — 50 мс, грудная мышца колибри — 8 мс). Из этого можно сделать вывод, что скорость движений когтя попугая зависит от высвобождения энергии, запасенной в упругом сухожилие, и от низкой инерции когтя. Столь быстрые движения когтей позволяют попугаям моментально приспосабливать свой захват в зависимости от геометрии и текстуры поверхности посадки.
Что касается степени захвата когтей, то она зависит и от диаметра жердочки, и от текстуры (3G). Самый выраженный захват наблюдается в случае диаметра 38 мм. Стоит уточнить, что речь идет именно о захвате когтей, а не всех лапок целиком. То есть, чем больше диаметр, тем больше нужно полагаться на когти, ибо лапки не помогут (3C, 3E). Угол когтя относительно поверхности сильно зависит от ее текстуры. К примеру, в случае более гладких поверхностей (тефлон) когти «загибаются» больше (3H), чем в случае шероховатой поверхности (наждачная бумага), так как в этом случае они могут просто чуть сильнее сжать поверхность, тем самым обеспечив надежное сцепление. Утрированно говоря, когти загибаются сильнее, когда поверхность сложнее сжать.

Изображение №4
Далее исследователи решили проверить, как с точки зрения механики изменяются показатели силы сжатия пальцами и изгиба когтей, и как они влияют на адаптацию птицы к той или иной поверхности во время посадки.
Тесты с различными текстурами жердочек показали, что и у подушечек пальцев (4А), и у когтей (4С) наблюдается схожая сдвигающая сила* в моменты скольжения и захвата.
Сдвигающая сила* — неравнозначные силы, толкающие одну часть тела в одном определенном направлении, а вторую часть тела — в противоположном.Для подушечек лап самым подходящим методом представления сдвигающей силы как функции нормальной силы для всех поверхностей является модель кулоновского трения (4B), позволяющая предсказать направление и величину силы трения между двумя телами, контактирующими сухими поверхностями (от того эту модель еще называют «модель сухого трения»).
Для когтей (4C) коэффициент трения может быть в 8 раз выше, чем у подушечек лап (4А). Такая разница может быть вызвана, помимо прочего, еще и ориентацией когтя относительно поверхности (4D): на самой мягкой поверхности сила трения остается относительно постоянной, но на грубой поверхности с увеличением угла когтя эта сила также увеличивается. Однако, даже при одинаковых углах измерения показывают, что коэффициент максимальной силы для когтей все равно выше, чем для подушечек (4E).
Чтобы компенсировать стохастический характер сцепления когтей, птицы могут воспользоваться распределением нагрузки и перемещением когтей по поверхности. Распределение нагрузки между несколькими когтями эффективно сужает распределение ожидаемой общей силы всех когтей (4Е). Кроме того, птицы могут вытягивать когти, что позволяет им повысить вероятность сцепления с неровной поверхностью, если таковая имеется. Это увеличивает ожидаемое значение силы трения (4F), что поясняет более выраженное загибание когтей на поверхностях, которые сложнее просто крепко сжать (3F). Вытягивание когтей дает преимущество в случае поверхностей, на которых неровностей мало, но они достаточно крупные. Однако это утверждение немного не согласуется с наблюдениями, ибо попугаи не шибко стремились вытягивать когти, а это говорит о том, что они не выбирают наилучшие неровности во время посадки. Следовательно, они загибают когти максимально, пока не будет достигнута достаточная сила для поддержания стабильного захвата. Если во время этого процесса данная сила не будет достигнута, то птица начнет корректировать положение когтей либо взлетит.

Изображение №5
Разница между коэффициентами трения для когтей и подушечек может еще объясняться геометрией этих частей тела и геометрией посадочной поверхности, с которой они контактируют. Подушечки обладают большей площадью и мягкой текстурой, что позволяет распределять нагрузку по многим точкам контакта, а это приводит к более равномерной силе трения. Когти, в свою очередь, грубые и обладают малой площадью, потому полагаются на прямое взаимодействие с геометрией поверхности, т.е. на зацепление за неровности.
Чтобы количественно оценить геометрические эффекты, ученые охарактеризовали форму когтя в сагиттальной плоскости по ширине как функцию высоты от кончика когтя (5А). В дополнение к использованию существующих поверхностных неровностей, когти могут также генерировать силы трения от поверхностной деформации.
Чтобы смоделировать это взаимодействие, были объединены измерения глубины проникновения когтя в различные поверхности (5B) и измерения геометрии самого когтя. Геометрия верхушки когтя была смоделирована сферой радиусом 50 мкм для нагрузок до 25% от массы тела (5С). Таким образом было установлено, что, например, глубина проникновения в жердочку из калифорнийского дуба составляет 10 мкм, т.е. 0.2% от длины внешней дуги когтя (примерно 5 мм).
Чтобы детальнее оценить влияние размера когтя на силу сцепления была создана еще одна модель с когтями разного диаметра верхушки, цепляющимися за разные поверхности (5D). Как и ожидалось, в случае отсутствия какого-либо проникновения когтей в поверхность более грубые поверхности являются более подходящими для успешного захвата (5Е).
Когда в модель были добавлены эффекты поверхностного проникновения (5F), было обнаружено, что полезная поверхность увеличивается. При этом чем выраженнее эффект, тем меньше угол, образованный между вертикальной осью и вектором чистой силы на когте. Если радиус и масса верхушки когтя масштабируются изометрически, а приложенные силы зависят от массы, то коэффициент глубины (глубина проникновения, деленная на радиус верхушки когтя) также будет масштабироваться изометрически.
Из этого можно сделать логический вывод, что мелким животным для эффективного сцепления с поверхностью достаточно мелких неровностей на этой поверхности.

Изображение №6
Объединив все вышеописанные модели для различных эффектов, сил и прочего, исследователи создали единую двумерную модель, в которой одновременно учитываются параметры когтей, лапок и тела в целом (6А).
Совокупность данных о механике контакта стоп и поверхности посадки позволили применить ограниченную оптимизацию для определения трехмерного «пространства захвата» птицы и необходимой комбинации силы и момента силы (6В). Было установлено, что важную роль в успешности посадки играет не только скорость птицы, но и направление: если вектор скорости птицы не направлен в центр жердочки, то птица может банально поскользнуться. Область стабильности захвата может активно расширяться за счет увеличения силы сжатия либо за счет самой текстуры поверхности, представляющей более доступное поверхностное трение (6C).
Чем меньше ветка, тем больше будет сила сжатия. Если пальцы птицы полностью охватывают ветку, то ее стабильность будет зависеть исключительно от силы мышц. Получается, что более тонкие ветки дают птицам больше шансов успешно сесть на них. Единственное исключение это гибкость: тонкие ветки могут прогибаться под весом птицы, отчего стабильность захвата может быть потеряна.
Оценка всех основополагающих сил, участвующих в процессе посадки птицы на жердочку разной геометрии и текстуры, выявила чрезвычайную важность поверхностного трения в формировании стабильного захвата. Однако, стоит отметить, что даже в случае скольжения по поверхности, далеко не всегда это происходит произвольно. Незначительное скольжение в момент контакта с поверхностью позволяет поглотить энергию во время приземления, быстрее обнаружить неровности с высоким коэффициентом трения.
Для более детального ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых.
Эпилог
Данный труд показал, что далеко не только полет является сложным процессом с точки зрения механики, аэродинамики и анатомии птиц. Процесс приземления также весьма непрост. Он объединяет в себе ряд физических сил, которые в правильной комбинации позволяют птице успешно и безопасно приземлиться. Зная это, по словам самих исследователей, человек сможет создать автономного летающего робота, которого не придется ловить в момент посадки или заранее подготавливать ему посадочную площадку. Птицы покорили небеса намного раньше нас, и нам есть чему у них поучиться. Кто-то может назвать это исследование нерациональной тратой человеко-часов, а кто-то — великим открытием. В любом случае, любые новые знания, будь то о квантово-классическом переходе или о кинематике когтей попугая во время посадки, важны. Если бы мы не имели жажды к новым знаниям, то до сих пор бы бегали с дубинками вокруг пещеры, утрировано говоря, конечно.
Пятничный офф-топ:
Попугаи не только отлично летают (и приземляются, как показало это исследование), но и веселиться умеют.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Попугаи не только отлично летают (и приземляются, как показало это исследование), но и веселиться умеют.
Благодарю за внимание, оставайтесь любопытствующими и отличных всем выходных, ребята! :)
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас оформив заказ или порекомендовав знакомым, 30% скидка для пользователей Хабра на уникальный аналог entry-level серверов, который был придуман нами для Вас:Вся правда о VPS (KVM) E5-2650 v4 (6 Cores) 10GB DDR4 240GB SSD 1Gbps от $20 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?